
Cómo la cisteína contribuye al cierre de estomas en respuesta a la sequía en plantas
El cierre de estomas es la más importante y evidente respuesta al estrés hídrico inducido por la sequía en plantas. Fundamental en este proceso es la activación de canales aniónicos lentos y rápidos (SLAC y QUAC, por sus siglas en inglés) modulados por el ácido abscísico (ABA) [1,2]. Estos canales facilitan la salida de Cl–, NO3– y SO42- desde las células de guarda provocando la despolarización del potencial de membrana hacia voltajes más positivos. La consecuencia de esta despolarización es la activación de canales rectificadores de K+ y el eflujo de este catión. Esto reduce el potencial osmótico lo que se traduce en la salida de agua y reducción del turgor de las células de guarda, y finalmente en el cierre del estoma [3].
Aun cuando por mucho tiempo se aceptó que la señal química que activa los canales iónicos para provocar el cierre de estomas en respuesta a la sequía es el ABA sintetizado en raíces y movilizado a través del xilema, este dogma comienza a cambiar cuando se demostró que las células de guarda también tienen la capacidad de sintetizar ABA y provocar este efecto de manera autónoma [4]. De este modo, el ABA producido en raíces no sería la única señal química de larga distancia que gatilla el cierre de estomas.
El sulfato como señal de larga distancia
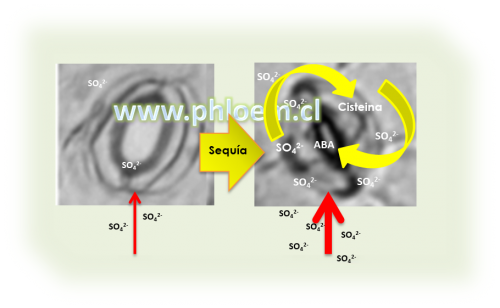
Numerosa evidencia indica que la adaptación de las plantas a la sequía está acompañada de incrementos en la absorción, concentración y flujo de sulfatos (SO42- ) en el xilema. De hecho, la sola aplicación de sulfatos gatilló todos los eventos necesarios y suficientes para inducir el cierre de estomas en la especie modelo Arabidopsis thaliana, de manera similar al efecto provocado por el ABA [1]. Uno de estos eventos es el aumento del peróxido de hidrógeno (H2O2) en las células de guarda, una especie de oxígeno reactivo cuya estimulación ocurre como consecuencia de la activación de la enzima NADPH oxidasa (Figura 1).

Si bien lo anterior indicaría que la participación del sulfato en la producción de H2O2 y consecuente cierre de estomas es independiente de ABA, la producción de H2O2 no ocurre en plantas mutantes deficientes en esta fitohormona demostrando que el cierre de estomas es más bien la consecuencia de la biosíntesis de ABA inducida por sulfatos.
La evidencia que asocia al sulfato con el cierre de estomas dependiente de ABA apunta al sulfuro (S2-), un metabolito intermediario en la asimilación del sulfato que se origina de la reducción del sulfito (SO32-) en una reacción catalizada por la enzima sulfito reductasa (SiR) [5] (Figura 1). Sin embargo, plantas mutantes deficientes en SiR son incapaces de producir H2O2 indicando que el compuesto azufrado asociado al cierre de estomas es sintetizado corriente abajo del sulfuro.
La conexión entre la síntesis de ABA y la cisteína
La última reacción en la biosíntesis de ABA en plantas es catalizada por la aldehído oxidasa (AO), una enzima que requiere del cofactor molibdeno (MoCo) sulfurado (MoCo-S) para que sea funcional. Como se mencionó en un artículo anterior [6], la adición de la molécula de azufre a MoCo para generar MoCo-S es catalizada por la enzima moco sulfurasa (ABA3), y esta molécula se origina de la cisteína (Figura 1).
En Arabidopsis, la sola aplicación de cisteína en hojas es suficiente para inducir el cierre de estomas de manera similar a la aplicación de sulfato, y elevar los niveles endógenos de ABA de manera similar a la aplicación de ABA. Esto demuestra que la asimilación del sulfato en la cisteína es una condición necesaria para su funcionalidad como señal química asociada al cierre de estomas dependiente de ABA en Arabidopsis y otras especies [1,2].
La relevancia biológica de la biosíntesis de ABA dependiente de cisteína se evaluó en plantas mutantes deficientes en cisteína, en suelos con baja disponibilidad hídrica [1]. En esta condición, las plantas desarrollaron el fenotipo característico del estrés por sequía demostrando que el sulfato puede gatillar el cierre de estomas a través de la inducción de la biosíntesis de ABA controlado por la cisteína.
Conclusión
Los estomas son una de las principales vías de comunicación de las plantas con el ambiente permitiendo la entrada de CO2 para realizar fotosíntesis y la salida de O2 como subproducto. Durante este intercambio gaseoso, los estomas quedan expuestos a la pérdida de agua por transpiración lo que puede ser fatal cuando ocurre en periodos de baja disponibilidad hídrica en el suelo. Es en este tipo de episodios donde las señales químicas que gatillan el cierre de estomas deben ser rápidamente activadas por el ABA.
El hecho que la sola aplicación de sulfatos, o de cisteína, sea suficiente para estimular la síntesis de ABA y provocar el cierre de estomas en respuesta a la sequía posicionan al azufre como una señal de comunicación raíz-parte aérea esencial en la adaptación de las plantas a uno de los estreses ambientales de mayor impacto en los cultivos. En esta señal, la molécula de azufre de la cisteína juega el papel más relevante.
PHLOEM, Agosto de 2019
www.phloem.cl
___________________________________________________________________________________________________________
Referencias
- Batool S, Uslu VV, Rajab H, Ahmad N, et al. 2018. Sulfate is incorporated into cysteine to trigger ABA production and stomatal closure. Plant Cell 30: 2973–2987.
- Malcheska F, Ahmad A, Batool S, Müller HM, et al. 2017. Drought-enhanced xylem sap sulfate closes stomata by affecting ALMT12 and guard cell ABA synthesis. Plant Physiol. 174: 798-814.
- Peñaloza E. 2018. La esencialidad del potasio en el funcionamiento de los estomas. Phloem, Diciembre 2017. www.phloem.cl.
- Bauer H, Ache P, Lautner S, Fromm J, et al. 2013. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr. Biol. 23: 53–57.
- Peñaloza E. 2019. El azufre y la tolerancia de las plantas al estrés ambiental: el papel de la cisteína. Phloem, Junio 2019. www.phloem.cl.
- Peñaloza E. 2018. El papel del molibdeno en la respuesta de las plantas al estrés oxidativo. Phloem, Febrero 2019. www.phloem.cl.