
Glucosinolatos; mecanismo natural de defensa en brásicas y su conexión con el azufre
Los glucosinolatos (GSL) son compuestos orgánicos característicos de la familia Brassicaceae reconocidos como pesticidas naturales en plantas y por sus propiedades anticancerígenas y antibióticas en el hombre. Estos compuestos derivan del metabolismo secundario y son sintetizados a partir de la glucosa y de aminoácidos. Su síntesis ocurre en todos los tejidos de una planta, aun cuando la concentración y diversidad de los GSL es generalmente mayor en raíces debido probablemente a una mayor y constante presión de pestes.
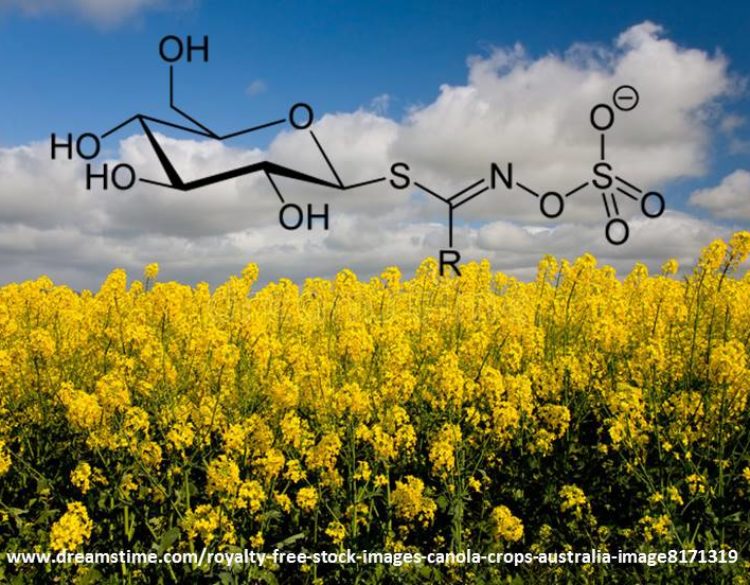
Para que cumplan su función, los GSL deben primeramente ser hidrolizados en una reacción catalizada por la enzima mirosinasa, una proteína que está presente en todas las especies que acumulan estos metabolitos. Una característica particular de los GSL y la enzima mirosinasa es que ambos se sintetizan y ubican en diferentes células de un mismo tejido. Esta separación espacial es propia de compuestos que participan en mecanismos naturales de defensa en plantas y, en brásicas, tiene por objetivo prevenir la hidrólisis de los GSL en compuestos que pudieran ser tóxicos para la célula, o cuando el mecanismo de defensa no es requerido por la planta.
El sistema glucosinolato-mirosinasa es activado cuando la célula pierde su integridad como consecuencia del ataque de insectos, herbívoros, patógenos, o cualquier tipo de daño mecánico. El daño físico a la célula permite el contacto entre el GSL y la enzima que lo hidroliza generando glucosa, sulfatos, y un intermediario inestable a partir del cual se producen numerosos compuestos con diversa actividad biológica [1]. Uno de estos compuestos son los isotiocianatos (Figura 1).

Los isotiocianatos y su potencial biofumigante
Los isotiocianatos (ITC) son compuestos volátiles derivados de la hidrólisis de GSL en raíces de crucíferas en descomposición o dañadas mecánicamente, y reconocidos como pesticidas naturales por sus efectos tóxicos o repelentes sobre plagas y patógenos. Este efecto fue descrito en raps (Brassica napus) por investigadores australianos en la década de los 90, quienes introdujeron el concepto de biofumigación para referirse a la acción biocida de los residuos de esta especie.
El compuesto responsable de la acción biocida de los ITC en raps se identificó como 2-feniletil isotiocianato (2-FE ITC). Este compuesto deriva de la degradación (hidrólisis) de gluconasturtiin, el GSL de mayor abundancia en raíces de raps canola y otras brásicas, y prácticamente ausente en la parte aérea. Adicionalmente, 2-FE ITC es el GSL de mayor toxicidad para patógenos del suelo, insectos y nemátodos, y su presencia en miembros de la familia Brassicaceae se ha vinculado al hecho que las especies de esta familia no se asocien con hongos micorricicos arbusculares [1,2,3].
La presencia de 2-FE ITC en raíces indica que es posible desarrollar genotipos de raps canola con alto potencial de biofumigación, sin comprometer la calidad nutricional de la semilla y de sus subproductos. Esto demuestra que la síntesis de GSL es órgano-específica y, paralelamente, explica la ausencia de correlación entre el contenido de GSL en la semilla y en la raíz de raps canola. En línea con esta tendencia, actualmente se está explorando la posibilidad de bloquear el transporte de GSL a la semilla en brásicas [4], lo que permitiría fortalecer el mecanismo natural de defensa de los GSL en la parte aérea de la planta.
La conexión entre los glucosinolatos y el azufre
Además de carbono, oxígeno y nitrógeno, la molécula de GSL contiene azufre (S) en su estructura cuya concentración puede alcanzar el 30% del S total en la planta. Si bien esto sugiere que los GSL pudieran funcionar como fuente de S utilizable en otros procesos metabólicos particularmente en condiciones de deficiencia de S, esto no necesariamente ocurre en la planta ya que el reciclaje de S desde los GSL está condicionado a la funcionalidad del sistema glucosinolato-mirosinasa (Figura 1).
Más que el reciclaje desde los GSL, recientemente se demostró que la deficiencia de S induce la expresión de genes cuyas proteínas reprimen la síntesis de GSL en brásicas [5] y que, en estas situaciones, la planta optimiza la utilización del S re-dirigiéndolo a la síntesis de proteínas en desmedro de la producción de GSL (Figura 2).

El sistema glucosinolato–mirosinasa en raps
La reconocida efectividad del raps como pre cultivo se ha atribuido en parte importante a la supresión de patógenos del suelo por GSL liberados desde la fitomasa en descomposición. Aun cuando este efecto biocida se ha demostrado en diversos patógenos del suelo como Gaeumannomyces, Rhizoctonia y Fusarium, la evidencia se origina principalmente de estudios in vitro o en condiciones controladas. En tanto, la información colectada de ambientes naturales es más restringida y de resultados menos predictivos por tratarse de procesos biológicos.
Entre los factores que pueden incidir sobre el funcionamiento de este sistema natural de defensa se incluyen la baja eficiencia de conversión de GSL a ITC en el suelo, la baja biomasa de raíces del raps y el tiempo de residencia de la fitomasa. Adicionalmente, y considerando que la degradación o hidrólisis de los GSL es altamente dependiente del tipo de suelo y su temperatura y humedad, la función biofumigante de los residuos debe entenderse como parte de un sistema integrado de control de pestes en donde los sulfatos (y el nitrógeno) desempeñan un papel importante.
PHLOEM, Julio de 2017
www.phloem.cl
Referencias
- Redonvikovic VR, Glivetic T, Delonga K, Vorkapic-Furac J. 2008. Glucosinolates and their potential role in plant. Period. Biol. 110: 297–309.
- Sarwar M, Kirkegaard JA, Wong PTW, Desmarchelier JM. 1998. Biofumigation potential of brassicas. III. In-vitro toxicity of isothiocyanates to soil-borne fungal pathogens. Plant Soil 201: 103-112.
- van Dam NM, Tytgat TOG, Kirkegaard JA. 2009. Root and shoot glucosinolates: a comparison of their diversity, function and interactions in natural and managed ecosystems. Phytochem. Rev. 8: 171–186.
- Nour-Eldin HH, Madsen SR, Engelen et al. 2017. Reduction of antinutritional glucosinolates in Brassica oilseeds by mutation of genes encoding transporters. Nat. Biotechnol.35: 377–382. doi:10.1038/nbt.3823.
- Aarabi F, Kusajima M, Thoge T, et al. 2016. Sulfur-defieciency-induced repressor proteins optimize glucosinolate biosynthesis in plants. Sci. Adv. 2(10):e1601087, eCollection; doi: 10.1126/sciadv.1601087.